



Ma perché abbiamo cinque dita?
Verso la soluzione del mistero dell'autopoiesi degli esseri viventi
“Sulle dita di una mano /
sono in cinque a far baccano”
Ops! La filastrocca non dice che a volte a far baccano sono in sei:

La polidattilia, in particolare quella con un dito in più sul lato del mignolo, è più comune di quanto si pensi, arrivando in alcune popolazioni fino a 1 su 150 nati. Nell’uomo è spesso dovuta a varianti del gene GLI3, e a volte del gene GLI1. Questi due geni, insieme a GLI2, formano una piccola “famiglia” genica, che produce dei fattori di trascrizione, cioè che regolano l’attività di altri geni.
Ma perché la “norma” è cinque dita, e come succede che un’alterazione di un singolo gene produca un intero dito supplementare?
In altre parole, come fa un arto in formazione a decidere che alla sua estremità si devono formare, in un tessuto indifferenziato, cinque e solo cinque (come direbbero i matematici) abbozzi di dita? E com’è che una semplice mutazione genetica riesce a “disturbare il manovratore”, inducendolo a programmare lo sviluppo di un dito in più?
Fino a non molti anni fa, a questa domanda si poteva solo rispondere che ci trovavamo davanti a uno dei misteri più affascinanti della biologia: il mistero della formazione delle strutture corporee, cioè di come in un organismo si generino forme complesse a partire da un tessuto omogeneo. Sul come ciò accada, l’embriologia ha rivelato dettagli anche molto precisi; ma quali sono i meccanismi, come emerga un ordine, un pattern, in un substrato multicellulare indifferenziato, restava una domanda senza risposta.
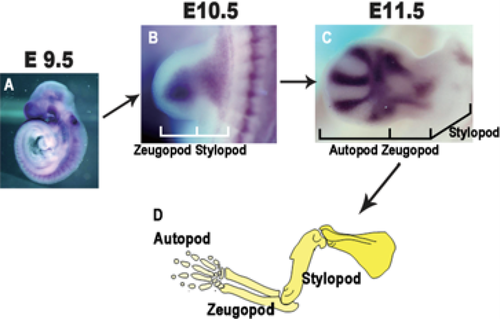
L’arto dei tetrapodi
L’arto pentadattilo (con cinque dita) è una caratteristica tipica dei vertebrati a quattro zampe, i tetrapodi terrestri e marini, e si riscontra in anfibi, rettili, uccelli e mammiferi. Si tratta di una struttura altamente conservata nell’evoluzione, che rappresenta un piano corporeo di base sorprendentemente stabile. Il fatto che l’adulto di molte specie abbia meno di cinque dita non deve trarre in inganno: nell’embrione di tutti i tetrapodi moderni si formano inizialmente cinque abbozzi digitali, che in alcune specie vengono poi modificati o riassorbiti nel corso dello sviluppo; nel cavallo addirittura ne rimane uno solo.
Nell’embrione dei tetrapodi, l’arto prende origine da una piccola gemma di cellule embrionali che si espande e si organizza progressivamente. A un certo punto dello sviluppo, nella porzione terminale dell’arto in formazione, le cellule del mesenchima iniziano a condensarsi in cinque regioni distinte: è qui che prendono forma gli abbozzi cartilaginei delle future ossa del cosiddetto autopodio (mano o piede).
Ma come si determina questo processo di condensazione, un vero e proprio arricchimento locale di cellule, che poi secernono cartilagine, ripetuto ad intervalli regolari? Negli esseri viventi non esiste un manovratore che diriga l’orchestra.
Vari articoli scientifici nell’ultimo ventennio hanno provato a trovare la risposta, fra cui questo (Sheth et al.):
I geni Hox regolano la strutturazione delle dita controllando la lunghezza d’onda di un meccanismo di tipo Turing
In combinazione con simulazioni al computer, i nostri risultati sostengono un meccanismo di tipo Turing alla base della formazione delle dita, in cui il numero dei geni Hox distali modula il periodo o la lunghezza d'onda.
Ohibò, che c’entra Alan Turing?

Alan Turing, oltre al suo lavoro pionieristico nell’informatica e nella crittografia, si interessò anche al mistero di cui si è parlato: come si formano spontaneamente le strutture regolari negli organismi viventi, tipo le strisce sul manto di una zebra o, appunto, le dita di una mano, che sono la versione 3D delle strisce. Nel 1952, pubblicò un articolo visionario (di ben 67 pagine) in cui propose che una configurazione disomogenea, ma regolare, di cellule o pigmenti potesse emergere spontaneamente da due o anche più sostanze che interagiscono e si diffondono a velocità diverse. Questo meccanismo, detto modello di reazione-diffusione, che è rimasto dimenticato nel cassetto della conoscenza umana per decenni, è stato riscoperto nel nuovo secolo, e oggi viene considerato un riferimento fondamentale per spiegare la formazione di strutture periodiche negli embrioni.
Il modello di reazione-diffusione di Turing
Dall’articolo del 1952:
Si suggerisce che un sistema di sostanze chimiche, dette morfògeni, che reagiscono tra loro e si diffondono attraverso un tessuto, sia adeguato a spiegare i principali fenomeni della morfogenesi. Tale sistema, per quanto inizialmente omogeneo, può sviluppare nel tempo un pattern o una struttura a causa di un'instabilità dell'equilibrio, innescata da perturbazioni casuali.
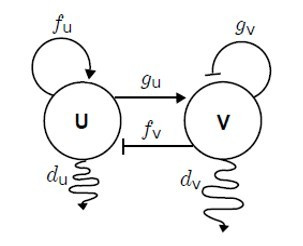
Nello schema di Turing, un composto u è un attivatore di se stesso (autocatalitico) e anche di un'altro composto v (frecce), il quale invece inibisce sia la propria sintesi che quella di u (linee tronche). I due composti si diffondono nell’ambiente circostante, v più velocemente di u (serpentine).
Il trattamento matematico di tale modello non è semplice. L’evoluzione temporale del sistema è descritto da un sistema di due equazioni differenziali accoppiate:
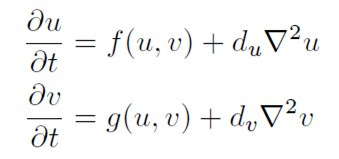
Turing dimostrò matematicamente che tale sistema di due sostanze chimiche in grado di reagire tra loro e di diffondersi nello spazio con velocità differenti può dare origine a “instabilità dinamiche” capaci di produrre strutturazione spaziale stabile e ripetitiva.
In altre parole, da una situazione iniziale omogenea – in cui le concentrazioni delle sostanze sono uniformi – può emergere spontaneamente nel tessuto una distribuzione spazialmente periodica, cioè un’alternanza regolare di massimi e minimi di concentrazione. In pratica si formano “onde stabili” di morfogeni, la cui lunghezza d’onda dipende dai valori dei parametri del modello, le quali alla fine determinano la configurazione delle strutture emergenti.
Turing non si limitò a formulare il modello teorico: in alcuni casi particolari, riuscì anche a calcolarne analiticamente le soluzioni, e a ottenere esempi numerici che mostravano la formazione delle onde stabili. Considerando che il calcolo delle derivate parziali e l’integrazione numerica di sistemi di equazioni differenziali accoppiate era nel 1950 un’impresa titanica, l’impegno intellettuale di Turing fu veramente considerevole.
Il modello di Turing e le dita dei topi
E come si articola il modello di Turing nella formazione delle dita dei topi?
Uno dei risultati più eclatanti mostrati nell’articolo di Sheth et al. è questo:
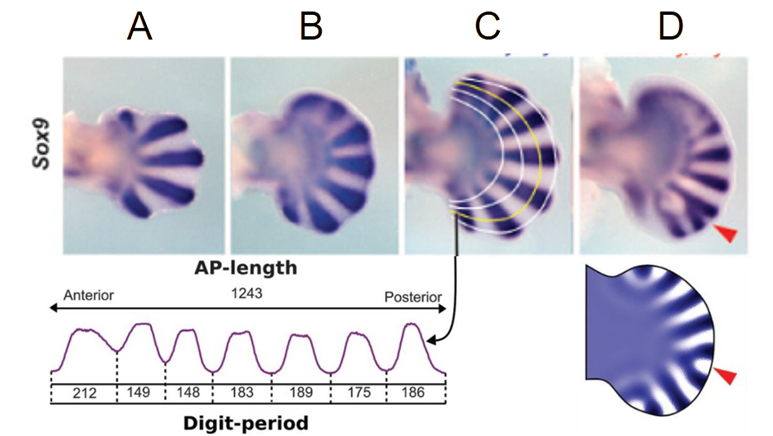
Le quattro fotografie mostrano la colorazione blu, al giorno 12.5, delle zone dell’autopodio dove è espresso il gene Sox9, che come abbiamo visto è coinvolto nella produzione della cartilagine.
A: autopodio di topo normale.
B: autopodio di un topo col gene Gli3 inattivato; analogamente a quanto accade nell’uomo si sviluppa polidattilia.
C: autopodio in un topo col gene Gli3 inattivato e con una delle due copie di un secondo gene, Hoxa13, inattivata. Il grafico derivato in basso mostra il dettaglio delle “onde” di colorazione Sox9 lungo il profilo tracciato dalle linee gialle. La prima onda a sinistra del grafico (corrispondente alla prima in alto nella foto) sembra indicare due primordi di dita fusi.
D: autopodio in un topo col gene Gli3 inattivato e con ambedue le copie del gene Hoxa13 inattivate. Il numero dei primordi delle dita aumenta e si disarticola fino quasi alla scomparsa delle onde, e compare anche una biforcazione (freccia rossa). La figura in basso è ottenuta con una simulazione al computer di un modello che impiega le condizioni minime per soddisfare le condizioni di instabilità di Turing, che riesce anche a riprodurre la biforcazione.
A conferma della validità del modello, il lavoro di Sheth et al. include un’analisi degli autopodi di topi Gli3-nulli che hanno da 0 a 4 copie inattive di due geni Hox correlati, il già visto Hoxa13 e il gene Hoxd11-13:

La prima foto a sinistra corrisponde alla situazione B della figura precedente: un topo con il gene Gli3 inattivo e con i due geni Hox normali (+/+, +/+); l’ultima foto a destra è invece di un topo con tutti i tre geni inattivi. Si osserva una progressiva polidattilia via via che aumenta il numero delle copie di geni Hox inattive (da 0 a 4), fino alla completa scomparsa della strutturazione. E’ notevole la corrispondenza con la simulazione al computer del modello di Turing facendo variare il valore di un singolo parametro.
Una linea di ricerca aperta
Il lavoro di Sheth et al. ha fornito una prova convincente dell’idea che un meccanismo di tipo Turing stia alla base della strutturazione delle dita, mostrando come mutazioni nei geni Hox possano modulare la lunghezza d'onda della concentrazione di morfogeni. Tuttavia, la loro simulazione numerica è più un'analogia, un modello matematico in grado di interpretare il fenomeno, senza però identificare i morfogeni reali.
In questo settore la ricerca è molto attiva. Un passo fondamentale è proprio l'identificazione dei componenti molecolari reali che agiscono come attivatori e inibitori. Ad esempio, un lavoro recente indica tre specifici morfogeni (Wnt, Sox9, e Bmp) come il cuore della rete di Turing che guida la formazione delle dita:

È indubbiamente sorprendente come, a decenni di distanza, un’intuizione geniale di Turing, guidata da un’analisi meticolosa e da un’idea semplice quanto ambiziosa,
Lo scopo di questo articolo è discutere un possibile meccanismo attraverso il quale i geni di uno zigote possano determinare la struttura anatomica dell’organismo che ne risulta,
si sia rivelata nell’era della genomica uno strumento potente per affrontare una delle questioni più profonde della biologia: l’origine della forma.
In questa prospettiva la forma finale di un organismo che si origina da una singola cellula (lo zigote), come il numero e la disposizione delle dita, ma così come di un’infinità di altre caratteristiche, non obbedisce ad un programma genetico che ne specifica i dettagli costruttivi: essa emerge invece da interazioni fra molecole diffusibili che agiscono come regolatori dell’attività dei geni, e la cui proprietà fondamentale è quella di retroagire in senso positivo o negativo sulla propria sintesi. In fondo questo è il meccanismo cardine dell’autopoiesi degli esseri viventi.